-
中文名稱:Recombinant Human Oxysterol-binding protein 1 (OSBP), partial
-
品名簡稱:Recombinant Human OSBP protein, partial
-
貨號:CSB-EP017243HU
-
說明書:
-
規格:¥1536
-
圖片:
-
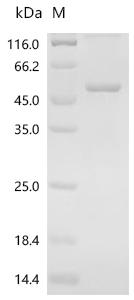 (Tris-Glycine gel) Discontinuous SDS-PAGE (reduced) with 5% enrichment gel and 15% separation gel.
(Tris-Glycine gel) Discontinuous SDS-PAGE (reduced) with 5% enrichment gel and 15% separation gel.
-
-
其他:
產品詳情
-
純度:Greater than 85% as determined by SDS-PAGE.
-
生物活性:Not Test
-
基因名:OSBP
-
Uniprot No.:
-
種屬:Homo sapiens (Human)
-
蛋白長度:Partial
-
來源:E.coli
-
分子量:50.7 kDa
-
表達區域:418-795aa
-
氨基酸序列NLWSIMKNCIGKELSKIPMPVNFNEPLSMLQRLTEDLEYHELLDRAAKCENSLEQLCYVAAFTVSSYSTTVFRTSKPFNPLLGETFELDRLEENGYRSLCEQVSHHPPAAAHHAESKNGWTLRQEIKITSKFRGKYLSIMPLGTIHCIFHATGHHYTWKKVTTTVHNIIVGKLWIDQSGEIDIVNHKTGDKCNLKFVPYSYFSRDVARKVTGEVTDPSGKVHFALLGTWDEKMECFKVQPVIGENGGDARQRGHEAEESRVMLWKRNPLPKNAENMYYFSELALTLNAWESGTAPTDSRLRPDQRLMENGRWDEANAEKQRLEEKQRLSRKKREAEAMKATEDGTPYDPYKALWFERKKDPVTKELTHIYRGEYWECK
Note: The complete sequence may include tag sequence, target protein sequence, linker sequence and extra sequence that is translated with the protein sequence for the purpose(s) of secretion, stability, solubility, etc.
If the exact amino acid sequence of this recombinant protein is critical to your application, please explicitly request the full and complete sequence of this protein before ordering. -
蛋白標簽:C-terminal 6xHis-tagged
-
產品提供形式:Liquid or Lyophilized powder
Note: We will preferentially ship the format that we have in stock, however, if you have any special requirement for the format, please remark your requirement when placing the order, we will prepare according to your demand. -
緩沖液:If the delivery form is liquid, the default storage buffer is Tris/PBS-based buffer, 5%-50% glycerol. If the delivery form is lyophilized powder, the buffer before lyophilization is Tris/PBS-based buffer, 6% Trehalose, pH 8.0.
-
復溶:We recommend that this vial be briefly centrifuged prior to opening to bring the contents to the bottom. Please reconstitute protein in deionized sterile water to a concentration of 0.1-1.0 mg/mL.We recommend to add 5-50% of glycerol (final concentration) and aliquot for long-term storage at -20°C/-80°C. Our default final concentration of glycerol is 50%. Customers could use it as reference.
-
儲存條件:Store at -20°C/-80°C upon receipt, aliquoting is necessary for mutiple use. Avoid repeated freeze-thaw cycles.
-
保質期:The shelf life is related to many factors, storage state, buffer ingredients, storage temperature and the stability of the protein itself.
Generally, the shelf life of liquid form is 6 months at -20°C/-80°C. The shelf life of lyophilized form is 12 months at -20°C/-80°C. -
貨期:3-7 business days
-
注意事項:Repeated freezing and thawing is not recommended. Store working aliquots at 4°C for up to one week.
-
Datasheet & COA:Please contact us to get it.
產品評價

相關產品
靶點詳情
-
功能:Lipid transporter involved in lipid countertransport between the Golgi complex and membranes of the endoplasmic reticulum: specifically exchanges sterol with phosphatidylinositol 4-phosphate (PI4P), delivering sterol to the Golgi in exchange for PI4P, which is degraded by the SAC1/SACM1L phosphatase in the endoplasmic reticulum. Binds cholesterol and a range of oxysterols including 25-hydroxycholesterol. Cholesterol binding promotes the formation of a complex with PP2A and a tyrosine phosphatase which dephosphorylates ERK1/2, whereas 25-hydroxycholesterol causes its disassembly. Regulates cholesterol efflux by decreasing ABCA1 stability.
-
基因功能參考文獻:
- the component proteins of the machinery, OSBP, VAP, SAC1, and PITPNB, are all essential host factors for AiV replication. Importantly, the machinery is directly recruited to the RNA replication sites through previously unknown interactions of VAP/OSBP/SAC1 with the AiV proteins and with ACBD3. PMID: 29367253
- results demonstrate that Sac1 expression in either the ER or Golgi apparatus has a minimal impact on the PI-4P that regulates OSBP activity or recruitment to contact sites PMID: 28471037
- Cholesterol transfer, PI4P consumption, and control of membrane lipid order by endogenous OSBP have been described. PMID: 28978670
- Data suggest that OSBP shifts the distribution of phosphatidylinositol 4-phosphate upon localization to endoplasmic reticulum-Golgi contact sites. PMID: 26601944
- Our results identify OspB as a regulator of mTORC1 and mTORC1-dependent cell proliferation early during S. flexneri infection and establish a role for IQGAP1 in mTORC1 signaling PMID: 26473364
- These results suggest that poliovirus proteins modulate PI4KB activity and provide PI4P for recruitment of OSBP to accumulate unesterified cholesterol on virus-induced membrane structure for formation of a virus replication complex. PMID: 24527995
- OSBP-mediated back transfer of phosphatidylinositol 4-phosphate might coordinate the transfer of other lipid species at the endoplasmic reticulum-Golgi interface. PMID: 24209621
- OSBP is required for efficient replication of intracellular S. Typhimurium. PMID: 21988961
- Data indicate that phosphorylation on two serine-rich motifs, S381-S391 (site 1) and S192, S195, S200 (site 2), specifically controls oxysterol-binding protein (OSBP) activity at the endoplasmic reticulum (ER). PMID: 22875984
- PKD negatively regulates HCV secretion/release by attenuating OSBP and CERT functions by phosphorylation inhibition. This study identifies the key role of the Golgi components in the HCV maturation process. PMID: 21285358
- Results identify a novel substrate of protein kinase D at the Golgi, the oxysterol-binding protein OSBP. PMID: 20444975
- This review summarizes recent evidence of sterol transfer activity by OSBP, suggesting seemingly disparate functions that could be the result of alterations in membrane sterol distribution or ancillary to this primary activity. PMID: 20545625
- Electrostatic interaction between oxysterol-binding protein and VAMP-associated protein A revealed by NMR and mutagenesis studies. PMID: 20178991
- OSBP was found to function as a cholesterol-binding scaffolding protein coordinating the activity of two phosphatases to control the extracellular signal-regulated kinase (ERK) signaling pathway PMID: 15746430
- Regulation of ceramide transport protein by OSBP, sterols, and vesicle-associated protein reveals a novel mechanism for integrating sterol regulatory signals with ceramide transport and phingomyelin synthesis in the Golgi apparatus. PMID: 16571669
- OSBP is able to sense both membrane cholesterol and oxidized sterols and link this information to the ERK1/2 signaling pathway. PMID: 18165705
- functional role of OSBP in the HCV maturation process. PMID: 19570870
顯示更多
收起更多
-
亞細胞定位:Cytoplasm, cytosol. Cytoplasm, perinuclear region. Golgi apparatus membrane; Peripheral membrane protein. Endoplasmic reticulum membrane; Peripheral membrane protein. Golgi apparatus, trans-Golgi network.
-
蛋白家族:OSBP family
-
組織特異性:Widely expressed.
-
數據庫鏈接:
Most popular with customers
-
Express system: Mammalian cell
Species: Homo sapiens (Human)
-
Express system: Mammalian cell
Species: Macaca fascicularis (Crab-eating macaque) (Cynomolgus monkey)
-
Recombinant Mouse Complement component C1q receptor (Cd93), partial (Active)
Express system: Mammalian cell
Species: Mus musculus (Mouse)
-
Recombinant Mouse Cytotoxic and regulatory T-cell molecule (Crtam), partial (Active)
Express system: Mammalian cell
Species: Mus musculus (Mouse)
-
Recombinant Human Dipeptidase 3(DPEP3), partial (Active)
Express system: Mammalian cell
Species: Homo sapiens (Human)
-
Recombinant Human Urokinase-type plasminogen activator(PLAU) (Active)
Express system: Mammalian cell
Species: Homo sapiens (Human)
-
Recombinant Human Tyrosine-protein kinase receptor UFO(AXL),partial (Active)
Express system: Mammalian cell
Species: Homo sapiens (Human)















