-
中文名稱:小鼠Mdm2重組蛋白
-
品名簡稱:Recombinant Mouse Mdm2 protein
-
貨號:CSB-YP013626MO
-
說明書:
-
規(guī)格:¥69
-
促銷:
-
圖片:
-
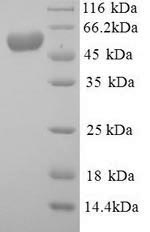 (Tris-Glycine gel) Discontinuous SDS-PAGE (reduced) with 5% enrichment gel and 15% separation gel.
(Tris-Glycine gel) Discontinuous SDS-PAGE (reduced) with 5% enrichment gel and 15% separation gel.
-
-
其他:
產(chǎn)品詳情
-
純度:Greater than 90% as determined by SDS-PAGE.
-
基因名:
-
Uniprot No.:
-
別名:Mdm2; E3 ubiquitin-protein ligase Mdm2; EC 2.3.2.27; Double minute 2 protein; Oncoprotein Mdm2; RING-type E3 ubiquitin transferase Mdm2; p53-binding protein Mdm2
-
種屬:Mus musculus (Mouse)
-
蛋白長度:Full Length
-
來源:Yeast
-
分子量:56.6kDa
-
表達(dá)區(qū)域:1-489aa
-
氨基酸序列MCNTNMSVSTEGAASTSQIPASEQETLVRPKPLLLKLLKSVGAQNDTYTMKEIIFYIGQYIMTKRLYDEKQQHIVYCSNDLLGDVFGVPSFSVKEHRKIYAMIYRNLVAVSQQDSGTSLSESRRQPEGGSDLKDPLQAPPEEKPSSSDLISRLSTSSRRRSISETEENTDELPGERHRKRRRSLSFDPSLGLCELREMCSGGSSSSSSSSSESTETPSHQDLDDGVSEHSGDCLDQDSVSDQFSVEFEVESLDSEDYSLSDEGHELSDEDDEVYRVTVYQTGESDTDSFEGDPEISLADYWKCTSCNEMNPPLPSHCKRCWTLRENWLPDDKGKDKVEISEKAKLENSAQAEEGLDVPDGKKLTENDAKEPCAEEDSEEKAEQTPLSQESDDYSQPSTSSSIVYSSQESVKELKEETQDKDESVESSFSLNAIEPCVICQGRPKNGCIVHGKTGHLMSCFTCAKKLKKRNKPCPVCRQPIQMIVLTYFN
Note: The complete sequence may include tag sequence, target protein sequence, linker sequence and extra sequence that is translated with the protein sequence for the purpose(s) of secretion, stability, solubility, etc.
If the exact amino acid sequence of this recombinant protein is critical to your application, please explicitly request the full and complete sequence of this protein before ordering. -
蛋白標(biāo)簽:N-terminal 6xHis-tagged
-
產(chǎn)品提供形式:Liquid or Lyophilized powder
Note: We will preferentially ship the format that we have in stock, however, if you have any special requirement for the format, please remark your requirement when placing the order, we will prepare according to your demand. -
緩沖液:If the delivery form is liquid, the default storage buffer is Tris/PBS-based buffer, 5%-50% glycerol.
Note: If you have any special requirement for the glycerol content, please remark when you place the order.
If the delivery form is lyophilized powder, the buffer before lyophilization is Tris/PBS-based buffer, 6% Trehalose. -
儲(chǔ)存條件:Store at -20°C/-80°C upon receipt, aliquoting is necessary for mutiple use. Avoid repeated freeze-thaw cycles.
-
保質(zhì)期:The shelf life is related to many factors, storage state, buffer ingredients, storage temperature and the stability of the protein itself.
Generally, the shelf life of liquid form is 6 months at -20°C/-80°C. The shelf life of lyophilized form is 12 months at -20°C/-80°C. -
貨期:Basically, we can dispatch the products out in 1-3 working days after receiving your orders. Delivery time may differ from different purchasing way or location, please kindly consult your local distributors for specific delivery time.Note: All of our proteins are default shipped with normal blue ice packs, if you request to ship with dry ice, please communicate with us in advance and extra fees will be charged.
-
注意事項(xiàng):Repeated freezing and thawing is not recommended. Store working aliquots at 4°C for up to one week.
-
Datasheet & COA:Please contact us to get it.
產(chǎn)品評價(jià)

相關(guān)產(chǎn)品
靶點(diǎn)詳情
-
功能:E3 ubiquitin-protein ligase that mediates ubiquitination of p53/TP53, leading to its degradation by the proteasome. Inhibits p53/TP53- and p73/TP73-mediated cell cycle arrest and apoptosis by binding its transcriptional activation domain. Also acts as a ubiquitin ligase E3 toward itself, ARRB1 and ARBB2. Permits the nuclear export of p53/TP53. Promotes proteasome-dependent ubiquitin-independent degradation of retinoblastoma RB1 protein. Inhibits DAXX-mediated apoptosis by inducing its ubiquitination and degradation. Component of the TRIM28/KAP1-MDM2-p53/TP53 complex involved in stabilizing p53/TP53. Also component of the TRIM28/KAP1-ERBB4-MDM2 complex which links growth factor and DNA damage response pathways. Mediates ubiquitination and subsequent proteasome degradation of DYRK2 in nucleus. Ubiquitinates IGF1R and SNAI1 and promotes them to proteasomal degradation. Ubiquitinates DCX, leading to DCX degradation and reduction of the dendritic spine density of olfactory bulb granule cells. Ubiquitinates DLG4, leading to proteasomal degradation of DLG4 which is required for AMPA receptor endocytosis. Negatively regulates NDUFS1, leading to decreased mitochondrial respiration, marked oxidative stress, and commitment to the mitochondrial pathway of apoptosis. Binds NDUFS1 leading to its cytosolic retention rather than mitochondrial localization resulting in decreased supercomplex assembly (interactions between complex I and complex III), decreased complex I activity, ROS production, and apoptosis.
-
基因功能參考文獻(xiàn):
- The proximal p53 inhibitor MDM2 is markedly downregulated in subcutaneous white and brown adipose tissues of mice during aging. Genetic disruption of MDM2 in adipocytes triggers canonical p53-mediated apoptotic and senescent programs, leading to age-dependent lipodystrophy and its associated metabolic disorders, including type 2 diabetes, nonalcoholic fatty liver disease, hyperlipidemia, and energy imbalance. PMID: 30131393
- To investigate whether MDM2C462A, which retains p53 binding, has p53-suppressing activity, we generated Mdm2C462A/C462A;p53ER/- mice. Adult Mdm2-null mice died approximately 7 days after tamoxifen-induced p53 activation, indicating that in the absence of MDM2, MDMX cannot suppress p53. p53 activity is higher in the presence of MDM2C462A than in the absence of MDM2. PMID: 29123033
- These results show that ischemic preconditioning increased neuronal MDM2 protein levels, which prevented ischemia-induced p53 stabilization and neuronal death. PMID: 29371613
- the disruption of Mdm2/p53 interaction affects the early-embryonic otic progenitor cells and their descendants. PMID: 28181574
- E2F6 suppresses Mdm2 expression in cells harboring the SNP309G allele but not the SNP309T allele. PMID: 28925402
- the MDM2-p53-PC signalling axis links mitochondrial metabolism to insulin secretion and glucose homeostasis, and could represent a therapeutic target in diabetes. PMID: 27265727
- Selective dysregulation of Mdm2 and Mdm4 alternative splicing underlies p53 anti-repression and motor neuron death in a mouse model of spinal muscular atrophy. PMID: 30012555
- Aging mouse models have revealed the complexity of the p53-Mdm2 axis and have solidly placed the p53 network as being key to many aspects of both pathological aging conditions and normal aging (review). PMID: 29192902
- The results indicated that simultaneously knocking down MDM2 and overexpressing p53 was able to inhibit proliferation and induce G1 cell cycle arrest in H1299 cells, compared with either alone. PMID: 29039579
- These results illustrate the importance of the cooperative activities of p53 and Mdm2 in a network of miRNAs that function to impose a barrier against aberrant cardiomyocyte cell cycle re-entry to maintain cardiac homeostasis. PMID: 28745540
- c-Abl phosphorylation of Mdm2 has a role in regulation of p53 tumor suppression and bone marrow failure PMID: 27956626
- Bre enhances osteoblastic differentiation by promoting the Mdm2-mediated degradation of p53. PMID: 28436570
- genetic and biochemical data support a role for Mdm2 in cardiac growth control through the regulation of p53, the Pgc-1 family of transcriptional coactivators and the pivotal antioxidant Pink1 PMID: 29267372
- The availability of large-scale genomic profiling datasets, like those from The Cancer Genome Atlas Research Network, have provided the opportunity to evaluate the consequences of MDM2 amplification and SNP inheritance across high-quality tumor samples from diverse cancer indications. [review] PMID: 27194168
- findings document contrasting effects of ATM-Mdm2 signaling on p53 tumor suppression and reveal that destabilizing Mdm2 by promoting its phosphorylation by ATM would be effective in treating oncogene-induced malignancies. PMID: 27568562
- the existence of an unusual functional interplay between STATs and CREB at the onset of adipogenesis through shared CRTC cofactors, is reported. PMID: 27362806
- Mdm2 expression is required for cell survival even in the absence of p53. Moreover, results suggest that p73 compensates for loss of p53. PMID: 28576884
- In Fmr1 KO neurons, Mdm2 is hyperphosphorylated, nuclear localized basally, and unaffected by MEF2 activation, which our data suggest due to an enhanced interaction with Eukaryotic Elongation Factor 1alpha (EF1alpha), whose protein levels are elevated in Fmr1 KO. Expression of a dephosphomimetic of Mdm2 rescues PSD-95 ubiquitination, degradation and synapse elimination in Fmr1 KO neurons. PMID: 28025327
- MDM2 is a non-redundant survival factor for proximal tubular cells by protecting them from spontaneous p53 overexpression-related cell death. PMID: 27882940
- The case emphasizes that MDM2 expression represents a possible pitfall in the diagnosis of spindle cell tumors. The differential diagnostic distinction between FDCS and a dedifferentiated liposarcoma is discussed. PMID: 27271257
- MDM2 is involved in fibroblast activation, mediating renal tubulointerstitial fibrosis via a p53-independent pathway dependant on Notch1 ubiquitination and proteasome degradation. PMID: 28100501
- These findings suggest that Mdm2 splice isoforms may play critical roles in the regulatory loop of p53/Mdm2-Mdm4 via a RING domain-mediated biochemical mechanism. PMID: 28166445
- both MDM2 and MDMX deletion-caused pancreatic defects are completely rescued by loss of p53, verifying the crucial role of the MDM2 and/or MDMX in regulating p53 in a spatio-temporal manner during the development, functional maintenance, and related disease progress of endocrine pancreas. PMID: 28118981
- Vif stabilization by CBFbeta is mainly caused by impairing MDM2-mediated degradation. PMID: 27758855
- These results demonstrated a critical prosurvival role for MDM2 in the oocytes PMID: 27912078
- we failed to detect any increase in p53 level in mutant oocytes, nor any other apoptotic marker, introgression of this targeted invalidation in p53-/- mice restored the fertility of females. This study is the first to show that Mdm2, but not Mdm4, has a critical role in oocyte survival and would be involved in premature ovarian insufficiency phenotype. PMID: 27364741
- Inhibition of MDM2 re-sensitizes rapamycin resistant renal cancer cells via the activation of p53. PMID: 27825169
- Inactivation of Mdm2 in sertoli cells triggers p53 activation and apoptosis as early as 15.5 days post conception with a significant increase in apoptosis. PMID: 26470726
- These findings elucidate a critical role of Mdm2-p53-Nedd4-2 signaling underlying the regulation of neural network synchrony and seizure susceptibility. PMID: 27000207
- These studies demonstrate that Mdm2 holds promise as a therapeutic target in combination with conventional therapy and may lead to new clinical therapies for Triple-negative breast cancers . PMID: 26494859
- there is a fine tuned balance in the interaction of ribosomal proteins with the MDM2/p53 axis which is important in normal hematopoiesis. PMID: 27042854
- Study showed that demonstrate that Mdm2 intertwines the mammalian target of RAPA, mTOR, and the receptor kinase GRK2 in regulating the desensitization/inactivation of the GPR17 receptor PMID: 26228571
- MDM2 mediates p73 ubiquitination PMID: 26025930
- MDM2 supports the PRC2 mediated repression of lineage-specific genes in stem cells, independent of p53. PMID: 26748827
- physiological activation of the 5S RNP-Mdm2-p53 pathway may contribute to functional decline of the hematopoietic system in a cell-autonomous manner over time. PMID: 25987256
- Ribosomal proteins L11 and L5 activate TAp73 by overcoming MDM2 inhibition. PMID: 25301064
- The Mdm2(SNP309-)(G) allele significantly impacts CRC through mechanisms outside the p53 pathway. PMID: 25435368
- MDM2 maintains homeostasis and long survival in podocytes preventing apoptosis. PMID: 25349197
- our results show MDM4-MDM2/p53-IGF1R as an original regulatory mechanism for CNS regeneration PMID: 25981963
- Mir-660 inhibits lung tumorigenesis by targeting MDM2-p53 interaction. PMID: 25501825
- L-GILZ stabilizes p53 proteins by decreasing p53 ubiquitination and increasing MDM2 ubiquitination. PMID: 25168242
- Interaction of BAI1 with the N-terminus (AA 1-200) of MDM2 in the brain modulates PSD-95 levels and thereby regulates synaptic plasticity. PMID: 25751059
- Mdm2regulates entry into myogenesis by targeting CEBPb for degradation by the 26 S proteasome. PMID: 25720496
- Src phosphorylation converts Mdm2 from a ubiquitinating to a neddylating E3 ligase. PMID: 25624478
- Data show that NEDD4-1 E3 ligase as a novel component of the tumor suppressor protein p53/proto-oncogene proteins c-mdm2 regulatory feedback loop that controls p53 activity during stress responses. PMID: 24413081
- Restoration of wild-type p53 expression in Mdm2-overexpressing tumors suppresses tumor growth. PMID: 24598047
- results reveal a novel p53- and Mdm2-independent oncogenic function of Mdmx that provides new insight into the many cancers that overexpress Mdmx. PMID: 24608433
- both MDM2 and MDMX are required for monitoring p53 activity during lens development, and they may function independently or synergistically to control p53 and maintain normal lens morphogenesis PMID: 25263199
- The regulation of Mdm2 by the E3 ubiquitin ligase APC/C is shown. It has important therapeutic implications for tumors with Mdm2 overexpression. PMID: 24804778
- TSLP induces mast cell development and aggravates allergic reactions through the activation of MDM2 and STAT6. PMID: 24751726
顯示更多
收起更多
-
亞細(xì)胞定位:Nucleus, nucleoplasm. Cytoplasm. Nucleus, nucleolus. Nucleus.
-
蛋白家族:MDM2/MDM4 family
-
組織特異性:Ubiquitously expressed at low-level throughout embryo development and in adult tissues. MDM2-p90 is much more abundant than MDM2-p76 in testis, brain, heart, and kidney, but in the thymus, spleen, and intestine, the levels of the MDM2 proteins are roughly
-
數(shù)據(jù)庫鏈接:
Most popular with customers
-
Recombinant Paguma larvata Angiotensin-converting enzyme 2 (ACE2), partial (Active)
Express system: Mammalian cell
Species: Paguma larvata (Masked palm civet)
-
Recombinant Human Heat-stable enterotoxin receptor (GUCY2C), partial (Active)
Express system: Mammalian cell
Species: Homo sapiens (Human)
-
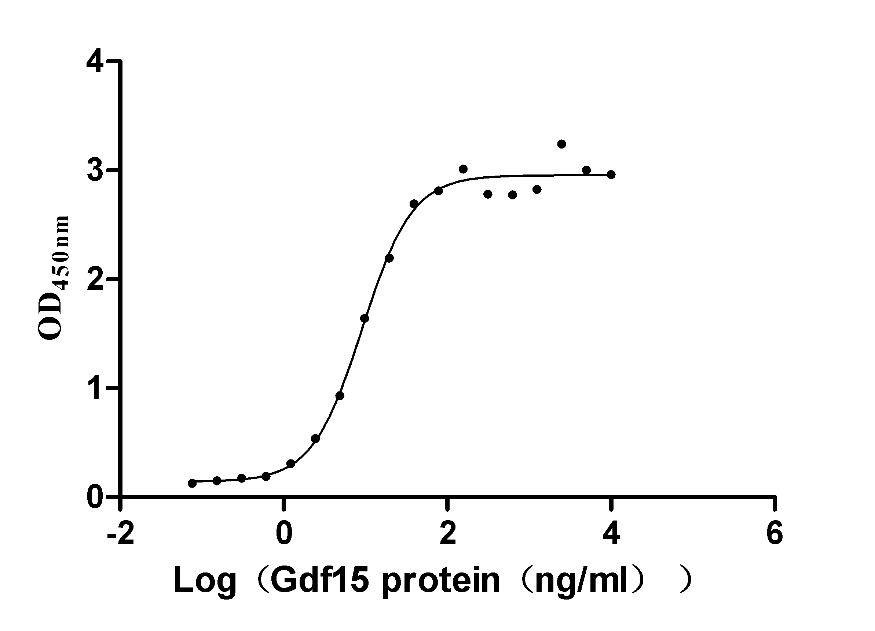
Recombinant Mouse GDNF family receptor alpha-like (Gfral), partial (Active)
Express system: Mammalian cell
Species: Mus musculus (Mouse)
-
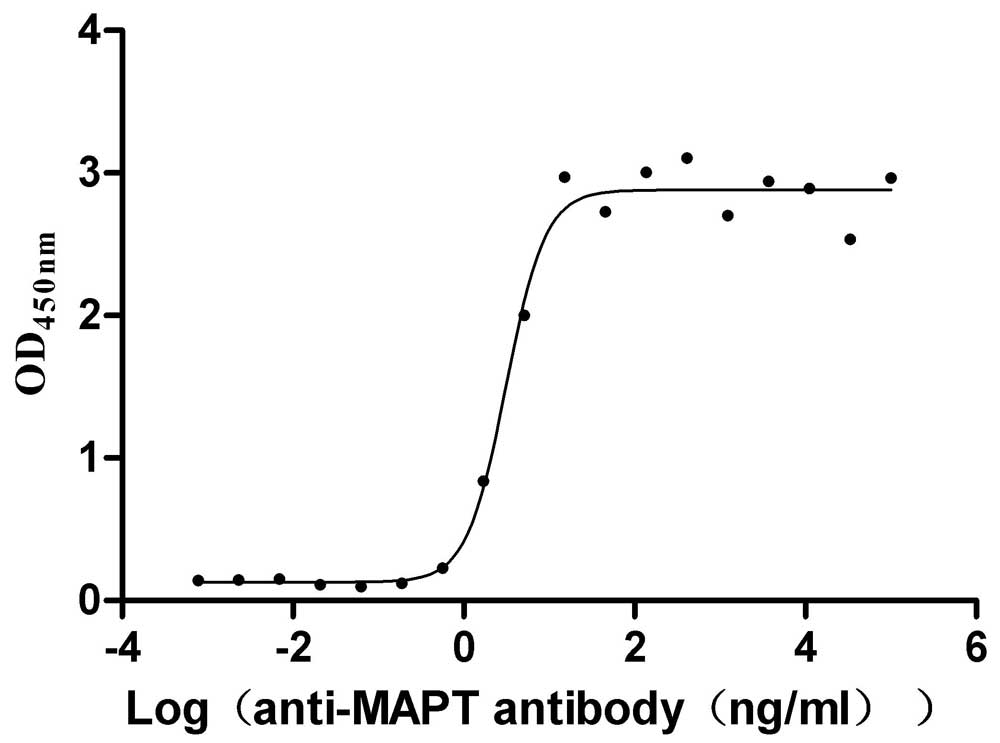
Recombinant Macaca mulatta Microtubule-associated protein tau (MAPT) (Active)
Express system: Mammalian cell
Species: Macaca mulatta (Rhesus macaque)
-
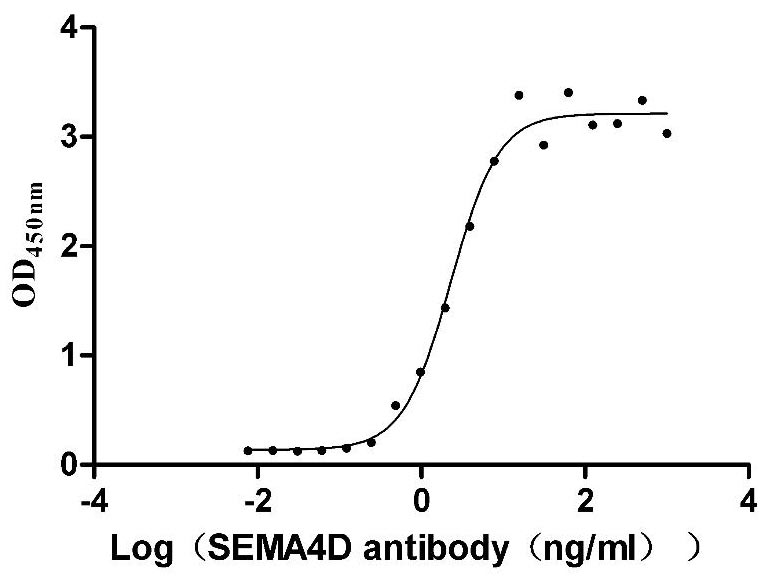
Recombinant Macaca mulatta Semaphorin-4D isoform 1 (SEMA4D), partial (Active)
Express system: Mammalian cell
Species: Macaca mulatta (Rhesus macaque)
-
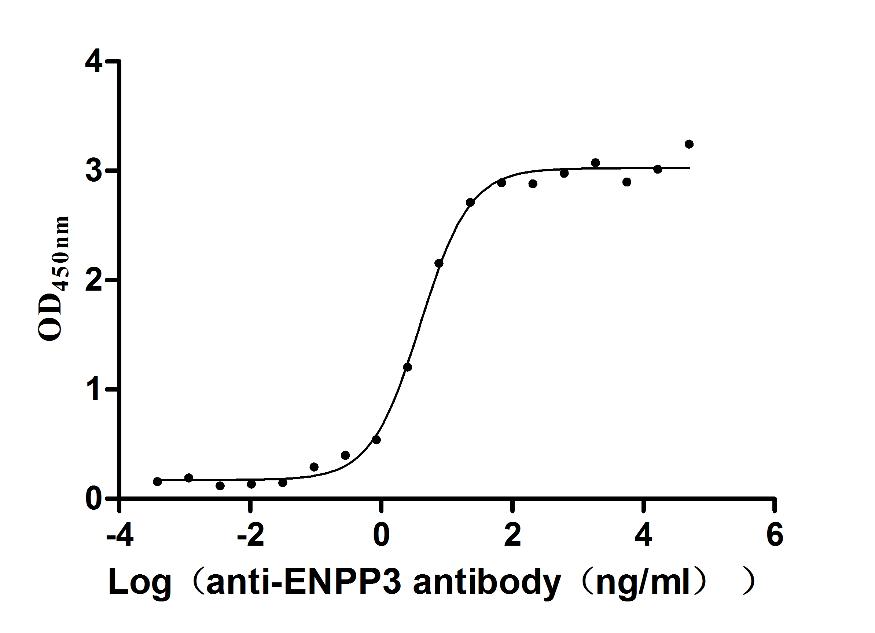
Express system: Mammalian cell
Species: Macaca fascicularis (Crab-eating macaque) (Cynomolgus monkey)
-
Recombinant Human Complement component C1q receptor (CD93), partial (Active)
Express system: Mammalian cell
Species: Homo sapiens (Human)
-
-AC1.jpg)
Recombinant Human Interleukin-17A (IL17A) (T26A) (Active)
Express system: Baculovirus
Species: Homo sapiens (Human)















