Recombinant Mouse Fibronectin type III domain-containing protein 5 (Fndc5), partial
In Stock Promotion-
中文名稱:小鼠Fndc5重組蛋白
-
品名簡稱:Recombinant Mouse Fndc5 protein, partial
-
貨號:CSB-EP815686MO
-
說明書:
-
規格:¥69
-
促銷:
-
圖片:
-
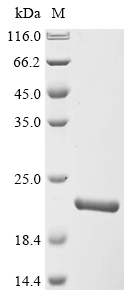 (Tris-Glycine gel) Discontinuous SDS-PAGE (reduced) with 5% enrichment gel and 15% separation gel.
(Tris-Glycine gel) Discontinuous SDS-PAGE (reduced) with 5% enrichment gel and 15% separation gel.
-
-
其他:
產品詳情
-
純度:Greater than 90% as determined by SDS-PAGE.
-
基因名:
-
Uniprot No.:
-
種屬:Mus musculus (Mouse)
-
蛋白長度:Partial
-
來源:E.coli
-
分子量:18.6 kDa
-
表達區域:29-140aa
-
氨基酸序列DSPSAPVNVTVRHLKANSAVVSWDVLEDEVVIGFAISQQKKDVRMLRFIQEVNTTTRSCALWDLEEDTEYIVHVQAISIQGQSPASEPVLFKTPREAEKMASKNKDEVTMKE
Note: The complete sequence may include tag sequence, target protein sequence, linker sequence and extra sequence that is translated with the protein sequence for the purpose(s) of secretion, stability, solubility, etc.
If the exact amino acid sequence of this recombinant protein is critical to your application, please explicitly request the full and complete sequence of this protein before ordering. -
蛋白標簽:N-terminal 6xHis-tagged
-
產品提供形式:Liquid or Lyophilized powder
Note: We will preferentially ship the format that we have in stock, however, if you have any special requirement for the format, please remark your requirement when placing the order, we will prepare according to your demand. -
緩沖液:If the delivery form is liquid, the default storage buffer is Tris/PBS-based buffer, 5%-50% glycerol. If the delivery form is lyophilized powder, the buffer before lyophilization is Tris/PBS-based buffer, 6% Trehalose.
-
復溶:We recommend that this vial be briefly centrifuged prior to opening to bring the contents to the bottom. Please reconstitute protein in deionized sterile water to a concentration of 0.1-1.0 mg/mL.We recommend to add 5-50% of glycerol (final concentration) and aliquot for long-term storage at -20°C/-80°C. Our default final concentration of glycerol is 50%. Customers could use it as reference.
-
儲存條件:Store at -20°C/-80°C upon receipt, aliquoting is necessary for mutiple use. Avoid repeated freeze-thaw cycles.
-
保質期:The shelf life is related to many factors, storage state, buffer ingredients, storage temperature and the stability of the protein itself.
Generally, the shelf life of liquid form is 6 months at -20°C/-80°C. The shelf life of lyophilized form is 12 months at -20°C/-80°C. -
貨期:3-7 business days
-
注意事項:Repeated freezing and thawing is not recommended. Store working aliquots at 4°C for up to one week.
-
Datasheet & COA:Please contact us to get it.
產品評價

相關產品
靶點詳情
-
功能:mediates beneficial effects of muscular exercise. Induces browning of white adipose tissue by stimulating UCP1 expression, at least in part, via the nuclear receptor PPARA.
-
基因功能參考文獻:
- A high-fat diet increased Fndc5 transcript levels in the skeletal muscle, but exercise had a minimal effect on the transcript level of Fndc5, whereas endurance training increased the protein content of FNDC5 in the skeletal muscle. PMID: 29365291
- These findings may lead to development of an Irisin-based therapy for elderly immobile osteoporotic and physically disable patients, and might represent a countermeasure for astronauts subjected to microgravity-induced bone and muscle losses. PMID: 28588307
- Glucocorticoid receptor positively regulates transcription of FNDC5 in the liver. PMID: 28240298
- Findings demonstrated that one bout of both uphill and downhill exercise trainings as well as 8 weeks of training could increase the expression of PGC-1alpha and FNDC5 genes in the muscle tissues and the UCP1 gene in the subcutaneous adipose tissue. PMID: 30218749
- Our study has indicated that irisin (FNDC5) possesses important anti-oxidant and anti-inflammatory properties and may protect cells from the damage induced by ROS under inflammatory conditions by activating of the Nrf2/HO-1 pathway. PMID: 29769428
- FNDC5 attenuated Oxidized low density lipoprotein-induced AMPK deactivation, hepatic stellate cells activation, connective tissue growth factor and transforming growth factor-beta upregulation and extracellular matrix deposition in mouse hepatic stellate cells. PMID: 30007970
- the coordinated expression of FNDC5 and PGC-1alpha may contribute to the increased levels of plasma irisin after exercise. PMID: 29415899
- Irisin(Fndc5) increases myogenic differentiation and myoblast fusion via activation of IL6 signaling. PMID: 29062100
- FNDC5 is overexpressed in skeletal muscle and adipose tissue of insulin resistant high fat-fed mice. PMID: 28676551
- This study for the first time showed that adipocytes are directly affected by irisin (Fndc5) and provides an evidence on anti-inflammatory action of irisin on fat cells. PMID: 28614774
- Mstn regulates Fndc5/Irisin expression and secretion through a novel miR-34a-dependent post-transcriptional mechanism; loss of Mstn in mice leads to the increased Fndc5/Irisin expression, which contributes to the browning of white adipocytes PMID: 27297797
- The secretion of FNDC5 from myotubes and beta-cells in response to exogenous fatty acids, the effects of recombinant FNDC5 on insulin biosynthesis and glucose-stimulated insulin secretion, and beta-cell apoptosis are reported. PMID: 28724742
- Irisin is upregulated in a murine model of fibrosis, but not in experimental NAFLD without significant fibrosis. PMID: 28472477
- Irisin improved endothelial function by modulating HO-1/ adiponectin axis in perivascular adipose tissue (PVAT) in HFD-induced obese mice. These findings suggest that regulating PVAT function may be a potential mechanism by which irisin improves endothelial function in obesity. PMID: 28595178
- The results indicate that FNDC5 deficiency impairs autophagy and FAO and enhances lipogenesis via the AMPK/mTOR pathway. FNDC5 deficiency aggravates whereas FNDC5 overexpression prevents the HFD-induced hyperlipemia, hepatic lipid accumulation, and impaired FAO and autophagy in the liver. PMID: 27504012
- This research is the first to show that irisin modulates macrophage activity by reducing reactive oxygen species (ROS) overproduction, which could suggest its potential anti-inflammatory properties. PMID: 28315350
- No FNDC5 expression was detected in normal or cancerous stomach tissues. FNDC5 expression in white and brown adipose tissues in the cancer group increased compared with the control and non-cancer groups. PMID: 27256459
- Report showed that irisin suppressed cholesterol synthesis in hepatocytes through the activation of 5' AMP-activated protein kinase (AMPK) and subsequent inhibition of transcription and nuclear translocation of SREBP2. PMID: 27211556
- PGC1alpha regulates FNDC5 and its processed and secreted peptide Irisin, which has been proposed to play a critical role in energy expenditure and to promote neural differentiation of mouse embryonic stem cells. Review. PMID: 26611102
- Irisin directly targets osteoblast, promoting osteoblast proliferation and differentiation via activating P38/ERK MAP kinase signaling cascades in vitro. PMID: 26738434
- irisin does not function through inflammatory NFKB activation like other myokines (such as TNFalpha). PMID: 26399516
- Muscle FNDC5 mRNA expression and irisin release are not IL-15-dependent in mice. PMID: 25920499
- irisin alleviates endothelial dysfunction in type 2 diabetes partially via reducing oxidative stresses through inhibiting signaling pathways suggesting that irisin may be a promising molecule for the treatment of vascular complications of diabetes. PMID: 26225842
- HFD induced obese mice showed decreased irisin secretion from adipose tissues, which might contribute to muscle insulin resistance. PMID: 26261526
- Our study also suggests the existence of irisin-specific receptor on the membrane of H9C2 cells. In conclusion, irisin in a certain concentration rage increased myocardial cell metabolism, inhibited cell proliferation and promoted cell differentiation PMID: 26305684
- In summary, these results suggested that the endothelium-dependent relaxation of irisin is mediated by the nitric oxide (NO)-guanosine 3', 5'-cyclic phosphate (cGMP)-dependent pathway. PMID: 26582714
- Fndc5 facilitates neural differentiation. This effect might be related to increased expression of BDNF following overexpression of Fndc5. PMID: 25572300
- This study used irisin radiolabeled with (125)I and small-animal SPECT/CT imaging to investigate the metabolic elimination and distribution of irisin in vivo. PMID: 25757030
- High intensity exercise results in increased expression of Fndc5 in skeletal muscle. PMID: 26166638
- Increase PGC-1alpha expression during cardiac precursor cells formation stage via rosiglitazone treatment increase FNDC5 and mitochondrial markers transcript levels which enhanced cardiac differentiation efficiency. PMID: 25936576
- The myokine irisin increases cortical bone mass and may be the molecular entity responsible for muscle-bone connectivity. PMID: 26374841
- findings shed light on the poorly understood regulation of irisin/FNDC5 by demonstrating a novel association between irisin and SMAD3 signaling in skeletal muscle PMID: 25648888
- Stage dependent expression of FNDC5 is affected by neural induction method used for neural differentiation. PMID: 24706335
- Data suggest that fibronectin type III domain-containing 5 protein (Fndc5) may be involved in cardiomyocyte differentiation. PMID: 25168892
- exists in muscle and serum of mice independent of exercise and is increased immediately after acute exercise PMID: 24644429
- FNDC5 (25 kDa) and irisin (12 kDa) were present in murine skeletal muscle and that irisin was circulating in murine serum. PMID: 24498244
- These findings link endurance exercise and the important metabolic mediators, PGC-1alpha and FNDC5, with BDNF expression in the brain. PMID: 24120943
- This immunohistochemical results demonstrate the expression of irisin immunoreactivity in three types of cells: skeletal, cardiac muscle, and Purkinje cells of the cerebellum. PMID: 23470775
- Fndc5 expression is required for the appropriate neural differentiation of mESCs. These data confirm the importance of Fndc5 in the generation and development of the nervous system. PMID: 23219938
- Within the muscle, Mstn(-/-) leads to increased expression of AMPK and its phosphorylation, which subsequently activates PGC1alpha and Fndc5. PMID: 23362117
顯示更多
收起更多
-
亞細胞定位:Cell membrane; Single-pass type I membrane protein. Peroxisome membrane; Single-pass type I membrane protein. Secreted. Note=Imported in peroxisomes through the PEX5 receptor pathway.; [Irisin]: Secreted. Note=Detected in the blood of individuals subjected to endurance exercise.
-
組織特異性:In adult, it is highly expressed in skeletal muscle, heart and brain.
-
數據庫鏈接:
Most popular with customers
-
Recombinant Macaca fascicularis Delta-like protein 3 (DLL3), partial (Active)
Express system: Mammalian cell
Species: Macaca fascicularis (Crab-eating macaque) (Cynomolgus monkey)
-
Recombinant Human Pancreatic adenocarcinoma up-regulated factor (ZG16B) (Active)
Express system: Mammalian cell
Species: Homo sapiens (Human)
-
-AC1.jpg)
Recombinant Mouse Claudin-18.2 (Cldn18.2)-VLPs (Active)
Express system: Mammalian cell
Species: Mus musculus (Mouse)
-
Recombinant Dog B-lymphocyte antigen CD20 (MS4A1)-VLPs (Active)
Express system: Mammalian cell
Species: Canis lupus familiaris (Dog) (Canis familiaris)
-
Recombinant Macaca fascicularis Trophoblast glycoprotein (TPBG), partial (Active)
Express system: Mammalian cell
Species: Macaca fascicularis (Crab-eating macaque) (Cynomolgus monkey)
-
Recombinant Mouse Cytotoxic and regulatory T-cell molecule (Crtam), partial (Active)
Express system: Mammalian cell
Species: Mus musculus (Mouse)
-
-AC1.jpg)
Recombinant Human Tumor necrosis factor ligand superfamily member 15(TNFSF15) (Active)
Express system: Mammalian cell
Species: Homo sapiens (Human)
-
Recombinant Human Tyrosine-protein kinase receptor UFO(AXL),partial (Active)
Express system: Mammalian cell
Species: Homo sapiens (Human)















