標簽蛋白對蛋白表達純化效率和蛋白活性有什么影響?
日期:2026-05-11 15:22:12
在原核、真核、哺乳動物細胞以及酵母、昆蟲細胞等重組蛋白表達體系中,標簽蛋白已經成為分子克隆與蛋白制備過程里必不可少的設計環節。所謂標簽蛋白,就是通過基因工程手段,在目的蛋白的 N 端或 C 端融合一段短肽或者完整功能蛋白結構,常見的包括 His標簽、GST標簽、MBP標簽、SUMO標簽、FLAG標簽、Strep標簽、Trx硫氧還蛋白標簽等。

2. 顯著改善蛋白可溶性,減少包涵體形成
3. 屏蔽蛋白酶酶切位點,減少胞內降解
4. 標簽帶來的表達負向影響
二、標簽蛋白對蛋白純化效率的影響
1. 實現特異性親和純化,大幅簡化工藝流程
3. 直接決定蛋白回收率與工藝穩定性
4. 標簽酶切去除帶來的純化附加影響
三、標簽蛋白對蛋白生物活性的影響
1. 改變蛋白空間構象,破壞天然折疊
加入標簽的初衷,本來是為了解決天然目的蛋白表達量低、可溶性差、難以純化、不易檢測等痛點,但在實際實驗和工業化生產中大家都會發現:標簽并不是萬能buff,它既能大幅提升表達量、簡化純化流程、提高回收率,也有可能改變蛋白空間構象、破壞活性中心、引發蛋白聚集、降低穩定性,甚至帶來免疫原性風險。
標簽蛋白就像一把雙刃劍,用得合適可以事半功倍,用得盲目則會出現表達很高、純化很純、但完全沒有生物活性的尷尬情況。本文就從蛋白表達效率、蛋白純化效率、蛋白生物活性三個核心維度,用通俗專業的語言,系統分析標簽蛋白帶來的正向影響、負面隱患以及不同標簽的適用規律。
一、標簽蛋白對蛋白表達效率的影響
重組蛋白表達的核心痛點主要集中在:整體表達量低、翻譯中途中斷、蛋白折疊錯誤、形成包涵體、胞內被蛋白酶降解等。標簽蛋白從翻譯起始、蛋白折疊、抗降解、可溶性維持四個層面,深刻改變目的蛋白的表達狀態。
1. 提升整體表達水平,優化翻譯起始效率
很多天然目的蛋白自身密碼子偏好性差、起始翻譯信號弱,單獨表達時核糖體結合效率低,整體表達量微乎其微。當在N端融合小分子標簽如6×His、FLAG、SUMO后,相當于給整個融合蛋白提供了強翻譯起始位點,能顯著提升核糖體識別與結合能力,讓宿主細胞更愿意啟動這條外源基因的翻譯過程。
尤其是原核大腸桿菌表達體系中,N端短肽標簽能夠穩定mRNA二級結構,避免轉錄翻譯提前終止,直接把目的蛋白的總表達量提升數倍甚至十幾倍。而像GST、MBP這類大分子標簽自身屬于高表達蛋白,融合后會帶動整條融合鏈高效轉錄翻譯,非常適合本身低表達的細胞因子、病毒蛋白、膜蛋白等。
2. 顯著改善蛋白可溶性,減少包涵體形成
這是標簽蛋白最大的實用價值之一。很多真核來源蛋白、膜蛋白、疏水蛋白、腫瘤抗原蛋白,在原核體系里極易出現疏水區域外露,分子間相互抱團,最終形成不溶于上清的包涵體。包涵體雖然蛋白總量很高,但都是錯誤折疊的無活性聚集體,復性難度大、成功率低,基本沒法用于后續功能實驗和藥物開發。
不同標簽的增溶能力差異非常明顯:MBP標簽增溶能力最強,堪稱難溶蛋白的 “救星”;SUMO、Trx、NusA標簽次之;GST標簽有較好增溶效果;6×His屬于弱增溶標簽,對本身可溶蛋白影響不大,但很難拯救極度疏水難溶的蛋白。
標簽本質上相當于給目的蛋白包裹了一層親水外殼,能夠掩蓋蛋白自身的疏水區域,減少分子間非特異性聚集,引導蛋白按照正確路徑折疊,大幅提高上清可溶性蛋白比例,這也是科研實驗中幾乎做難溶蛋白必加大分子助溶標簽的核心原因。
3. 屏蔽蛋白酶酶切位點,減少胞內降解
宿主細胞內部存在大量內源蛋白酶,外源重組蛋白很容易被識別為異源物質,被逐步剪切降解,最終只能檢測到碎片化蛋白,得不到完整全長目的蛋白。
標簽融合在蛋白末端,可以物理性遮擋目的蛋白末端的敏感酶切位點,同時改變整體蛋白表面電荷與空間結構,讓蛋白酶難以識別和結合,起到天然保護作用。像His、SUMO、GST這類標簽,都有很好的抗降解效果,能極大提高完整蛋白的占比,降低碎片化程度。
4. 標簽帶來的表達負向影響
凡事有利必有弊,標簽并不是加了就一定更好。首先,大分子標簽會增加翻譯負擔,MBP、GST這類幾十kDa的大標簽,會拉長翻譯時長、消耗宿主氨基酸與能量,部分情況下反而會導致宿主代謝壓力過大,整體表達量不升反降。
其次,標簽插入位置不合理也會破壞表達。多數蛋白適合N端標簽,如果目的蛋白C端是功能核心區、分泌信號區、跨膜結構區,強行加C端標簽會直接破壞信號識別,導致蛋白無法正常表達、分泌和定位。另外部分標簽自身電荷偏強,會改變整體蛋白等電點,引發胞內異常聚集,反而降低可溶性。
二、標簽蛋白對蛋白純化效率的影響
重組蛋白純化的核心訴求是:步驟少、純度高、回收率高、成本低、工藝可放大、條件溫和不損傷蛋白。標簽蛋白最大的工業價值,就是把傳統繁瑣的離子交換、疏水層析、凝膠過濾多步純化,簡化為親和層析一步純化,徹底改變了蛋白純化的邏輯。
1. 實現特異性親和純化,大幅簡化工藝流程
沒有標簽的天然蛋白,只能依靠分子大小、電荷、疏水性等理化性質進行分離,需要多步層析串聯,流程長、損耗大、純度有限、條件苛刻,還容易丟失活性。
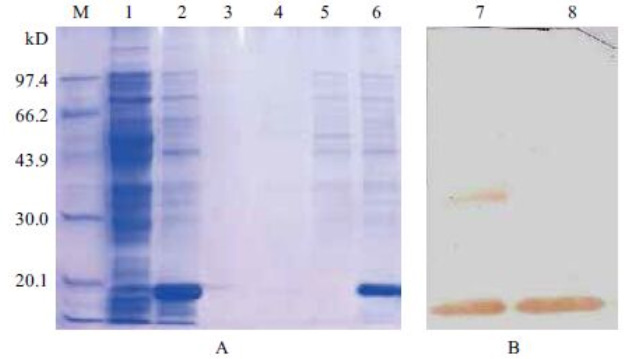
而帶標簽的融合蛋白,可以利用標簽與特異性填料的專屬結合實現一步抓取:His標簽結合鎳柱、GST結合谷胱甘肽填料、MBP結合麥芽糖樹脂、Strep標簽結合生物素填料、FLAG標簽結合特異性抗體填料。這種親和結合具有高度專一性,宿主雜蛋白基本不會非特異性吸附,一步洗脫就能得到高純度蛋白,極大縮短純化時間、減少操作步驟。
對于實驗室科研來說,節省大量實驗時間;對于工業化生產來說,簡化工藝流程意味著降低設備投入、減少人工成本、更容易工藝放大。
2. 影響純化純度與非特異性吸附
不同標簽的純化純度差異很大。Strep、FLAG標簽特異性極強,非特異性吸附極低,純化產物純度極高,適合對純度要求嚴苛的結構生物學、酶活實驗;GST、MBP標簽特異性也較好,純度穩定;而 6×His 標簽最大優勢是成本低、通用性強,但鎳柱容易和宿主內源組氨酸富集蛋白發生非特異性結合,在表達量偏低時,純度會略低于其他標簽,需要通過優化洗脫梯度、增加清洗步驟來改善。
同時,標簽大小也會影響層析柱載量和傳質效率,大分子標簽會增大整體流體力學半徑,擴散速度變慢,上樣流速不能太快,會降低純化批次效率;小分子標簽則無此問題,更適合大規模工業連續化生產。
3. 直接決定蛋白回收率與工藝穩定性
標簽和填料的結合強弱直接影響回收率。結合過弱,上樣過程中大量蛋白流穿,回收率偏低;結合過強,洗脫條件過于劇烈,容易導致蛋白變性、聚集、活性丟失。6×His標簽的優勢就是結合強度適中,洗脫條件溫和,咪唑洗脫不會破壞蛋白空間結構,回收率穩定,適配絕大多數活性蛋白制備;而GST、MBP結合力偏強,洗脫需要高濃度配體,成本更高,部分敏感蛋白容易在洗脫過程中失活。
另外從工業應用看,His標簽耐受強堿清洗、填料可反復再生使用,適配GMP生產和長期穩定工藝;而FLAG、Strep填料價格昂貴、耐堿差,只能用于科研小量制備,無法大規模產業化。
4. 標簽酶切去除帶來的純化附加影響
多數藥用蛋白、功能活性蛋白,不能保留外源標簽,需要設計TEV、凝血酶、PreScission等酶切位點,純化后切掉標簽。標簽的大小、酶切位點設計是否合理,直接影響酶切效率和后續二次純化難度。小分子His標簽酶切簡單、殘留少;大分子標簽容易出現空間位阻,導致酶切不完全,殘留融合蛋白,進一步增加純化負擔。
三、標簽蛋白對蛋白生物活性的影響
這是實驗中最容易被忽視、也最容易踩坑的環節。很多人只關注表達量和純化純度,忽略標簽對蛋白構象、活性中心、分子互作的影響,最后得到 “高表達、高純度、零活性” 的無效蛋白。標簽對活性的影響主要體現在空間構象、活性位點遮擋、蛋白聚集、穩定性及免疫原性五個方面。
1. 改變蛋白空間構象,破壞天然折疊
蛋白的生物活性完全依賴正確的三維空間構象。小分子短肽標簽如His、FLAG體積小、結構簡單,柔性強,對整體蛋白折疊干擾極小,絕大多數情況下不影響天然構象和生物活性;而GST、MBP這類大分子標簽自身擁有獨立球狀結構,融合后會改變目的蛋白整體空間排布,拉扯、扭曲原有折疊方式,導致構象異常、局部結構松散,直接喪失生理活性。
尤其對于酶類、細胞因子、受體胞外段、抗原蛋白,結構高度精密,一點點構象擾動都會讓底物結合、受體結合、抗原識別能力大幅下降。
2. 遮擋活性中心與分子互作位點
很多蛋白的活性位點、受體結合位點、酶催化中心位于蛋白末端區域,如果標簽剛好融合在活性端,會直接產生空間位阻,遮擋功能位點,導致蛋白無法和底物、受體、抗體發生正常結合,活性顯著降低甚至完全喪失。
這也是為什么做活性實驗時,優先切除標簽的核心原因。不切標簽測出來的酶活、結合活性、細胞活性數據,往往是失真的,無法代表天然蛋白真實功能。
3. 誘導蛋白聚集,降低均一性
部分標簽自身電荷偏高、疏水性較強,融合后會改變整個融合蛋白的表面電荷分布和疏水分布,破壞蛋白溶解性平衡,即使在上清中也容易形成二聚體、多聚體甚至可溶性聚集體。聚集體蛋白不僅生物活性大幅下降,還會在體內引發免疫清除、副作用升高,完全不符合藥用標準。
小分子標簽一般不會引發聚集,而大分子標簽如果位點設計不當,聚集概率會大幅上升,需要通過緩沖液pH、鹽離子濃度、溫度條件額外優化,增加實驗工作量。
4. 影響蛋白體內外穩定性與半衰期
合適的標簽可以提升蛋白熱穩定性、凍融穩定性、儲存穩定性,比如SUMO、Trx、MBP能夠穩定蛋白結構,降低降解速率,延長保存時間;但部分標簽會讓蛋白結構變得松散,熱穩定性變差,常溫、4℃放置極易降解失活,反復凍融后活性斷崖式下跌。
在體內應用層面,標簽會改變蛋白分子量、電荷、疏水性,影響體內循環半衰期、組織分布和清除速率,保留外源標簽的重組蛋白在動物體內很容易被免疫系統識別清除,藥效大打折扣。
5. 帶來免疫原性風險,限制藥用開發
科研實驗中可以暫時保留標簽,但臨床藥用重組蛋白、抗體融合蛋白絕對不能帶標簽。標簽屬于異源外源序列,進入人體后會被免疫系統識別為非自身物質,誘發抗體產生、炎癥反應、過敏反應,甚至引發嚴重免疫排斥。
同時標簽殘留還會影響藥物藥效、體內代謝和安全性評價,所以藥物研發中,標簽只作為純化工具,最終必須完全切除并去除標簽殘留,這也是工業上優先選用可酶切小標簽的根本原因。
四、不同常用標簽的綜合特性總結
從表達、純化、活性三個維度綜合來看:
6×His標簽:表達適配性廣、弱助溶、純化成本最低、工藝易放大、對活性干擾極小,是科研和工業最通用的首選標簽,唯一短板是純度略低于高端標簽。
GST標簽:增溶強、純化純度高,但分子量大、易干擾活性、洗脫成本高、不適合藥用大規模生產。
MBP標簽:增溶能力最強,專門拯救難溶膜蛋白和疏水蛋白,但體積過大、極易影響蛋白活性,僅適合科研前期獲取可溶性蛋白。
SUMO/Trx標簽:兼具助溶、穩構象、易酶切的優勢,對活性影響較小,適合毒性蛋白、難表達真核蛋白。
GST標簽:增溶強、純化純度高,但分子量大、易干擾活性、洗脫成本高、不適合藥用大規模生產。
MBP標簽:增溶能力最強,專門拯救難溶膜蛋白和疏水蛋白,但體積過大、極易影響蛋白活性,僅適合科研前期獲取可溶性蛋白。
SUMO/Trx標簽:兼具助溶、穩構象、易酶切的優勢,對活性影響較小,適合毒性蛋白、難表達真核蛋白。
FLAG/Strep標簽:純度極高、幾乎不影響活性,但填料昂貴、載量低、無法工業化放大,僅適用于小量高精度實驗。
標簽蛋白對蛋白表達、純化、活性的影響是全方位且辯證的。在表達層面,合理選擇標簽能夠提升轉錄翻譯效率、增強可溶性、減少包涵體、抵抗胞內降解;標簽過大或位點錯誤則會增加代謝負擔、降低表達水平。在純化層面,標簽實現了親和一步純化,簡化流程、提升純度和回收率,讓蛋白制備從復雜多步變成標準化流程,但標簽類型也決定了純化成本、工藝可放大性與產品純度上限。在活性層面,小分子標簽基本不干擾構象與功能,大分子標簽極易遮擋活性位點、改變空間構象、誘發聚集、降低穩定性,還存在明顯免疫原性風險。
實際實驗和生產中,選擇標簽的核心原則始終是:能選小標簽不選大標簽,能N端不隨便放C端,功能活性實驗盡量切除標簽,工業化生產優先His可酶切體系,難溶蛋白再選用MBP、SUMO等助溶標簽。只有合理搭配標簽類型、融合位置、酶切設計,才能既保證表達純化效率,又最大程度保留蛋白天然生物活性。
上一篇: 畢赤酵母表達系統重組蛋白表達與純化
下一篇: 最后一頁















