FGF2:從細胞增殖到腫瘤耐藥,一文讀懂這個多效性生長因子的臨床轉(zhuǎn)化潛力
日期:2026-03-27 15:45:55
成纖維細胞生長因子2(FGF2)是一種多功能生長因子,廣泛參與細胞增殖、血管生成、組織修復(fù)及腫瘤進展等過程。本文系統(tǒng)綜述了FGF2的分子結(jié)構(gòu)、信號通路(如PI3K/AKT/mTOR、MAPK/ERK)及其在腫瘤、神經(jīng)系統(tǒng)疾病、視網(wǎng)膜病變和骨骼疾病中的作用機制,旨在為FGF2相關(guān)疾病的機制研究與精準治療提供參考。
1. FGF2概述及其生物學(xué)意義
成纖維細胞生長因子2(FGF2),又稱堿性成纖維細胞生長因子(bFGF),是FGF家族的重要成員,在細胞增殖、分化、遷移、血管生成和組織修復(fù)等過程中發(fā)揮關(guān)鍵作用。FGF家族屬于結(jié)構(gòu)較為保守的肽類生長因子,主要通過與成纖維細胞生長因子受體(FGFRs)及肝素硫酸(HS)蛋白聚糖結(jié)合,激活下游信號通路并調(diào)控細胞行為 [1, 2]。研究表明,F(xiàn)GF2可促進FGFR構(gòu)象改變并增強其激活水平,而HS及其修飾在FGF2/FGFR信號調(diào)控中也具有重要作用 [1-3]。
FGF2參與多種關(guān)鍵生物學(xué)過程。在細胞增殖方面,YAP可通過上調(diào)FGFR表達增強神經(jīng)干細胞對FGF2的響應(yīng);在ER+乳腺癌中,F(xiàn)GF2還能激活ERα和PR信號,促進腫瘤細胞增殖并誘導(dǎo)內(nèi)分泌治療耐藥 [4, 5]。在血管生成方面,F(xiàn)GF2是重要的促血管生成因子,常與VEGF等分子共同參與血管形成與重塑,并在心肌損傷修復(fù)、視網(wǎng)膜病變和缺血性卒中等過程中發(fā)揮作用 [6-10]。在分化調(diào)控中,F(xiàn)GF2與成骨、成牙本質(zhì)分化密切相關(guān),同時在組織修復(fù)和抗纖維化中也表現(xiàn)出積極作用 [11-15]。
FGF2還存在低分子量(LMW)和高分子量(HMW)等不同亞型,兩者在細胞內(nèi)定位和功能上存在差異,并可能在疾病過程中發(fā)揮不同作用 [16]。同時,F(xiàn)GF2異常表達或信號失衡與多種疾病密切相關(guān),尤其涉及腫瘤進展、治療耐藥及部分炎癥相關(guān)疾病 [17-21]。總體來看,F(xiàn)GF2兼具重要的基礎(chǔ)研究價值和臨床轉(zhuǎn)化潛力,深入研究其作用機制、信號通路及疾病關(guān)聯(lián),對于理解相關(guān)疾病發(fā)生發(fā)展并探索新的治療策略具有重要意義。
2. FGF2的分子生物學(xué)特性與作用機制
2.1 FGF2的結(jié)構(gòu)、亞型與非經(jīng)典分泌
FGF2是FGF家族中功能多樣的重要成員,其蛋白結(jié)構(gòu)具有家族共同特征,即高度保守的中央β-折疊桶結(jié)構(gòu),該結(jié)構(gòu)域?qū)ζ渑cFGFR及硫酸肝素相關(guān)分子的結(jié)合至關(guān)重要。FGF2的生物學(xué)復(fù)雜性在很大程度上來源于其多種蛋白亞型。Fgf2基因可通過不同起始密碼子翻譯生成低分子量(約18 kDa)和高分子量(約22--24 kDa)異構(gòu)體 [16, 23]。其中,LMW FGF2主要定位于細胞質(zhì)和細胞核,而HMW FGF2則更多集中于細胞核。不同亞型在細胞內(nèi)定位和功能上的差異,使FGF2在不同組織和疾病背景下呈現(xiàn)出更復(fù)雜的調(diào)控模式。
值得注意的是,F(xiàn)GF2缺乏經(jīng)典信號肽,因此其分泌并不依賴傳統(tǒng)的內(nèi)質(zhì)網(wǎng)-高爾基體途徑,而是通過非經(jīng)典分泌方式釋放到細胞外。這一特性決定了FGF2在細胞外基質(zhì)中的沉積、局部濃度形成及旁分泌/自分泌調(diào)控均具有獨特性,也為其在發(fā)育、修復(fù)和疾病中的時空特異性作用提供了基礎(chǔ)。
2.2 FGF2與受體(FGFRs)及共受體(HSPGs)的結(jié)合與活化
FGF2信號轉(zhuǎn)導(dǎo)始于其與細胞表面FGFRs的特異性結(jié)合,這一過程通常依賴肝素硫酸蛋白聚糖(HSPGs)的協(xié)同作用。FGF2、FGFR和HSPG共同形成功能性三元復(fù)合物,被認為是FGF2發(fā)揮生物學(xué)活性的基礎(chǔ)。FGF2與受體結(jié)合后可促進FGFR二聚化,并引發(fā)胞內(nèi)酪氨酸激酶結(jié)構(gòu)域的磷酸化,繼而啟動一系列下游信號級聯(lián)反應(yīng)。
不過,F(xiàn)GF2與HS/HSPGs的作用并非簡單的"有或無"關(guān)系。已有研究顯示,降低FGF2與HS的結(jié)合能力并不會完全消除FGFR1信號 [2],說明共受體的貢獻可能具有情境依賴性。與此同時,特定類型的HS修飾,如2-O硫酸化,則對FGF2/FGFR1/Erk1/2通路的有效激活仍然至關(guān)重要 [3]。因此,F(xiàn)GF2受體復(fù)合體的形成與激活并不是單一分子事件,而是受到配體、受體及糖胺聚糖微環(huán)境共同調(diào)控的精細過程。
2.3 FGF2調(diào)控的細胞生物學(xué)過程
FGF2參與調(diào)控多種關(guān)鍵細胞行為,包括細胞增殖、遷移、分化、抗凋亡及血管生成等。在干細胞維持與增殖方面,F(xiàn)GF2對于骨髓間充質(zhì)干細胞等細胞群體的干性維持具有重要意義;在神經(jīng)系統(tǒng)中,YAP通過增強細胞對FGF2的響應(yīng)促進神經(jīng)干細胞擴增 [4]。在腫瘤環(huán)境中,F(xiàn)GF2則可推動腫瘤細胞增殖并促進治療耐受形成 [5, 17]。
在分化方面,F(xiàn)GF2對成骨和成牙本質(zhì)分化的調(diào)控尤其突出。它既可通過ERK-Runx2軸增強成骨分化 [13],也可在特定細胞背景下通過抑制PI3K/AKT通路促進SCAP成骨/成牙本質(zhì)分化 [11]。在組織修復(fù)和血管生成中,F(xiàn)GF2可通過與VEGF等因子協(xié)同,增強血管新生、改善局部供血,并促進損傷組織修復(fù) [6, 8, 10, 14]。因此,F(xiàn)GF2并非僅僅是單一功能生長因子,而是貫穿多類細胞過程的重要調(diào)控節(jié)點。
3. FGF2信號轉(zhuǎn)導(dǎo)通路與調(diào)控網(wǎng)絡(luò)
3.1 經(jīng)典的PI3K/AKT/mTOR信號通路
FGF2通過與FGFR結(jié)合激活多條細胞內(nèi)信號通路,其中PI3K/AKT/mTOR通路是其核心下游之一。FGFR激活后可招募并激活PI3K,促使PIP2轉(zhuǎn)化為PIP3,從而進一步激活A(yù)KT。活化的AKT可調(diào)節(jié)mTOR及其他下游效應(yīng)分子,最終影響細胞生長、代謝、存活和抗凋亡等過程。
在不同疾病和組織背景下,F(xiàn)GF2對PI3K/AKT/mTOR通路的調(diào)控具有多樣性。例如,在病理性血管生成中,PIK3IP1下調(diào)可增強PI3K-AKT信號,并提升FGF2等促血管生成因子的表達,從而加重異常血管生成 [9]。在組織修復(fù)中,CXCR4/Akt/FGF2軸可促進FGF2分泌并增強組織再生 [14]。但在某些分化場景中,F(xiàn)GF2又可能通過抑制PI3K/AKT通路發(fā)揮促分化作用,如在SCAP成骨/成牙本質(zhì)分化中所見 [11]。這說明FGF2與PI3K/AKT/mTOR通路之間并非單向激活關(guān)系,而是具有明顯的細胞類型和生物學(xué)情境依賴性。
3.2 MAPK/ERK及其相關(guān)信號通路
MAPK/ERK通路是FGF2介導(dǎo)細胞效應(yīng)的另一條經(jīng)典通路。FGF2與FGFR結(jié)合后,通過適配蛋白和Ras-Raf-MEK級聯(lián)反應(yīng)激活ERK,進而調(diào)控細胞生長、分化、遷移和應(yīng)激反應(yīng)。YAP可通過上調(diào)FGFR1-4表達增強細胞對FGF2的響應(yīng),并進一步激活MAPK和AKT信號,從而促進胚胎神經(jīng)干細胞增殖 [4]。在骨形成過程中,F(xiàn)GF2激活ERK后可增強Runx2蛋白穩(wěn)定性和轉(zhuǎn)錄活性,對成骨細胞分化和骨骼形成發(fā)揮積極作用 [13]。
然而,MAPK/ERK通路也是FGF2參與腫瘤耐藥的重要機制之一。在KIT突變型黑色素瘤中,F(xiàn)GF2軸通過MAPK通路抑制BIM表達,促進對奈洛替尼的耐藥 [18]。在膠質(zhì)瘤中,放療誘導(dǎo)的YAP激活可上調(diào)FGF2并進一步增強MAPK-ERK信號,形成放射抵抗 [19]。因此,F(xiàn)GF2-MAPK/ERK軸既是正常生理調(diào)控的重要通路,也是腫瘤進展和治療失敗的重要分子基礎(chǔ)。
3.3 FGF2信號的交叉對話與復(fù)雜調(diào)控機制
FGF2信號網(wǎng)絡(luò)并非獨立運行,而是與VEGF/VEGFR、Src、YAP、CXCR4等多條信號通路發(fā)生復(fù)雜交叉調(diào)控。以血管生成為例,F(xiàn)GF2與VEGF常共同參與血管形成與重塑,在缺血修復(fù)和視網(wǎng)膜病變中均表現(xiàn)出明顯協(xié)同作用 [6, 7, 8, 10]。在病理性環(huán)境中,Src通路激活可伴隨FGF2及其受體表達升高,增強促血管生成效應(yīng) [8]。此外,YAP既可通過上調(diào)FGFR增強細胞對FGF2的反應(yīng),也可在放療后促進FGF2分泌,從而形成促存活和耐藥環(huán)路 [4, 19]。
這種交叉調(diào)控進一步說明,F(xiàn)GF2并不是單一通路中的線性因子,而是多層級調(diào)控網(wǎng)絡(luò)中的關(guān)鍵樞紐。也正因如此,針對FGF2的治療策略通常難以通過單一環(huán)節(jié)實現(xiàn)理想效果,聯(lián)合阻斷多個節(jié)點往往更具潛力。
4. FGF2與相關(guān)疾病的病理生理學(xué)
4.1 FGF2在腫瘤發(fā)生發(fā)展及耐藥性中的作用
FGF2在多種腫瘤的發(fā)生發(fā)展中具有重要促癌作用,其機制涉及細胞增殖、遷移、侵襲、血管生成以及多種治療耐藥。ER+乳腺癌中,F(xiàn)GF2可通過激活雌激素非依賴性ER信號促進腫瘤生長,并導(dǎo)致內(nèi)分泌治療耐藥 [5, 17]。在KIT突變型黑色素瘤中,F(xiàn)GF2軸參與奈洛替尼耐藥形成,而MEK抑制劑可部分逆轉(zhuǎn)該過程 [18]。在非小細胞肺癌、乳腺癌和黑色素瘤中,F(xiàn)GF2/FGFR1高表達與TKI治療后的DTP形成相關(guān),提示FGFR信號代償性激活是腫瘤獲得耐藥的重要機制之一。
除耐藥外,F(xiàn)GF2還與腫瘤侵襲性密切相關(guān)。其全身水平在部分胃癌和食管癌患者中升高,可能反映更強的疾病進展傾向 [20]。在膠質(zhì)瘤中,F(xiàn)GF2參與放療抵抗形成,使其成為潛在的放療增敏靶點 [19]。因此,F(xiàn)GF2不僅是腫瘤生長相關(guān)因子,更是影響治療反應(yīng)和臨床結(jié)局的重要分子。
4.2 FGF2在神經(jīng)系統(tǒng)疾病與視網(wǎng)膜病變中的作用
FGF2在神經(jīng)系統(tǒng)中兼具促進增殖、營養(yǎng)支持和組織保護等多重作用。研究顯示,YAP可通過增強神經(jīng)干細胞對FGF2的響應(yīng)來促進其增殖 [4]。此外,F(xiàn)GF2在帕金森病模型中表現(xiàn)出神經(jīng)營養(yǎng)和神經(jīng)保護作用,可支持多巴胺能神經(jīng)元存活,并減輕MPP+或魚藤酮誘導(dǎo)的神經(jīng)損傷。
在視網(wǎng)膜病變中,F(xiàn)GF2既參與組織修復(fù),也與病理性血管生成密切相關(guān)。缺氧條件下,YY1乳酸化可增強FGF2轉(zhuǎn)錄并促進血管新生 [7];PIK3IP1下調(diào)和miRNA-15a/16-1簇缺失也均可通過不同機制提升FGF2相關(guān)信號,推動異常血管生成 [8,9]。與此同時,A1-PEG則可通過提升FGF2表達,在抑制病理性新生血管形成的同時促進血管修復(fù)和視網(wǎng)膜功能改善 [10]。這表明FGF2在神經(jīng)和視網(wǎng)膜疾病中具有明顯的雙重性,其作用方向高度依賴具體病理背景。
4.3 FGF2與骨骼、牙齒及結(jié)締組織疾病
FGF2在骨骼形成、重塑和維持中發(fā)揮重要作用。其通過激活ERK MAP激酶增強Runx2穩(wěn)定性和轉(zhuǎn)錄活性,從而促進成骨細胞分化和骨骼形態(tài)發(fā)生 [13]。在牙齒相關(guān)組織中,F(xiàn)GF2還能增強SCAP的成骨/成牙本質(zhì)分化能力,這為牙髓-牙本質(zhì)修復(fù)研究提供了重要依據(jù) [11]。
另一方面,F(xiàn)GF2信號失衡也與骨代謝異常相關(guān)。例如,苯甲酸鈉可通過抑制FGF2/p38/RUNX2通路導(dǎo)致成骨受阻和骨量流失 [12]。此外,F(xiàn)GF2在結(jié)締組織修復(fù)和抗纖維化中也具有重要意義,其衍生短肽P5可在肺纖維化模型中減輕EMT和膠原沉積 [15]。因此,F(xiàn)GF2在骨骼、牙齒及結(jié)締組織疾病中既是病理變化的重要調(diào)節(jié)因子,也是潛在干預(yù)靶點。
4.4 FGF2在炎癥、感染、心血管及代謝性疾病中的作用
FGF2在炎癥、感染、心血管和代謝性疾病中的作用同樣復(fù)雜。在心血管損傷修復(fù)中,F(xiàn)GF2與VEGF等因子共同參與血管新生,可改善梗死心肌供血并促進功能恢復(fù) [6]。在放射性腸損傷中,CXCR4/Akt/FGF2軸有助于促進腸道再生 [14]。在心肌梗死模型中,不同F(xiàn)GF2亞型對長期心功能和血管生成的影響并不一致,提示亞型差異可能影響疾病結(jié)局 [16]。
在炎癥和感染相關(guān)疾病中,F(xiàn)GF2的作用也逐漸受到關(guān)注。蛋白質(zhì)組學(xué)研究提示FGF2與牙周炎存在因果關(guān)聯(lián) [21]。在代謝相關(guān)病理性血管生成中,如糖尿病視網(wǎng)膜病變早期,F(xiàn)GF2與VEGF等因子共同參與病變進展。總體而言,F(xiàn)GF2在這些疾病中的作用并非單純有益或有害,而是具有明顯的環(huán)境依賴性和階段依賴性。
5. FGF2靶向藥物研究進展
目前靶向bFGF及相關(guān)信號通路的在研藥物類型多樣,包括Fc融合蛋白、小分子化藥及基因療法等,適應(yīng)癥覆蓋糖尿病性黃斑水腫、肝細胞癌、胰腺癌及脫髓鞘疾病等多個領(lǐng)域,研發(fā)階段從申請上市到臨床1期不等,涉及榮昌生物、基亞生物科技及深圳未知君生物等多家機構(gòu)。部分在研管線列舉如下:
| 藥物 | 靶點 | 藥物類型 | 在研適應(yīng)癥 | 在研機構(gòu) | 最高研發(fā)階段 |
|---|---|---|---|---|---|
| 伊立芙普α | VEGF x bFGF | Fc融合蛋白 | 糖尿病性黃斑水腫 | 濕性年齡相關(guān)性黃斑變性 | 榮昌生物制藥(煙臺)股份有限公司 | 申請上市 |
| 磺酸化單磷酸甘露寡聚糖 | VEGF-A x aFGF x bFGF | 聚合物 | 肝細胞癌 | 基亞生物科技股份有限公司 | 臨床3期 |
| AUP1602-C | M-CSF x bFGF | 工程菌 | 糖尿病足潰瘍 | 糖尿病足 | 靜脈曲張潰瘍 | 褥瘡 | 深圳未知君生物科技有限公司 | Aurealis Oy | Aurealis Therapeutics AG | 臨床2期 |
| Curcumin C3 complex | NF-κB x bFGF x mTOR | 小分子化藥 | 宮頸上皮內(nèi)瘤樣病變 | 人瘤病毒相關(guān)性宮頸癌 | 皮膚鱗狀細胞癌 | Emory University | Lsu Health Shreveport | 臨床2期 |
| Umedaptanib pegol | bFGF | 適配體 | 軟骨發(fā)育不全 | 糖尿病性黃斑水腫 | Ribomic, Inc. | 臨床2期 |
| Oleandrin | AP-1 x FLI1 x Na/K-ATPase x RPS6KB1 x bFGF x mTOR | 小分子化藥 | 胰腺癌 | 腫瘤 | U.S. Army Medical Research Institute for Infectious Diseases | Nerium Biotechnology, Inc. | The University of Texas MD Anderson Cancer Center | 臨床2期 |
| 綠原酸 | Akt x DNMT1 x LAG3 x SLC37A4 x TGF-β1 x bFGF | 小分子化藥 | 復(fù)發(fā)性膠質(zhì)瘤 | 晚期肺腺癌 | 晚期小細胞肺癌 | 晚期鱗狀細胞肺癌 | 腎腫瘤 | 肝癌等 | 四川九章生物科技有限公司 | 湖南中醫(yī)藥大學(xué) | Universiti Putra Malaysia | Brawijaya University | 臨床2期 |
| 成纖維細胞生長因子2基因療法(Id Pharma Co., Ltd.) | bFGF | 基因療法 | 間歇性跛行 | 外周動脈疾病 | 圍手術(shù)期缺血 | Kyushu University | Id Pharma Co., Ltd. | 臨床2期 |
| rHIgM22 | PDGFRs x bFGF | 單克隆抗體 | 脫髓鞘疾病 | Mayo Clinic | 臨床1期 |
(數(shù)據(jù)截止到2026年3月19日,來源于synapse)
6. FGF2研究工具
FGF2是一種具有廣泛生物學(xué)功能的多效性生長因子,在細胞增殖、分化、遷移、血管生成和組織修復(fù)等方面發(fā)揮核心作用。其通過與FGFRs結(jié)合,激活MAPK/ERK、PI3K/AKT以及其他多條信號通路,形成復(fù)雜而精細的調(diào)控網(wǎng)絡(luò)。與此同時,F(xiàn)GF2異常表達或信號失調(diào)又與多種疾病密切相關(guān),尤其在腫瘤進展、治療耐藥、視網(wǎng)膜病變、神經(jīng)系統(tǒng)疾病、骨代謝異常和纖維化等領(lǐng)域表現(xiàn)突出。CUSABIO提供FGF2重組蛋白、抗體及ELISA試劑盒產(chǎn)品,助力您進行相關(guān)機制研究及靶向藥物開發(fā)。
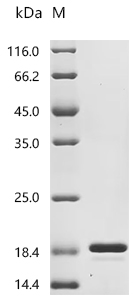
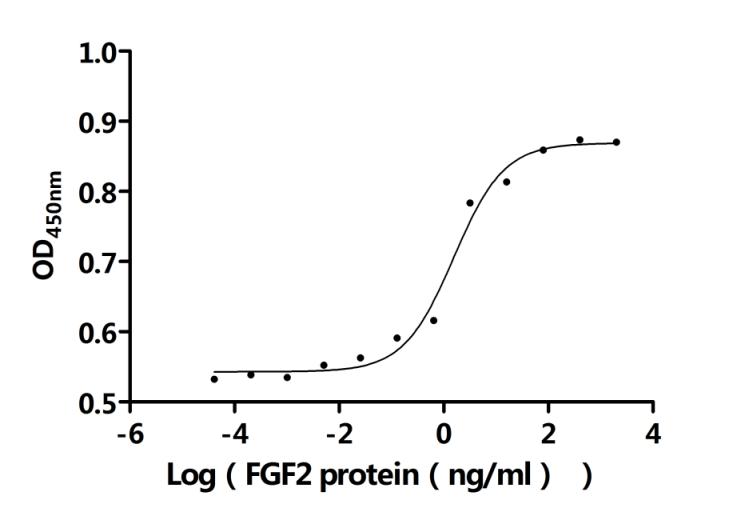
● FGF2 重組蛋白
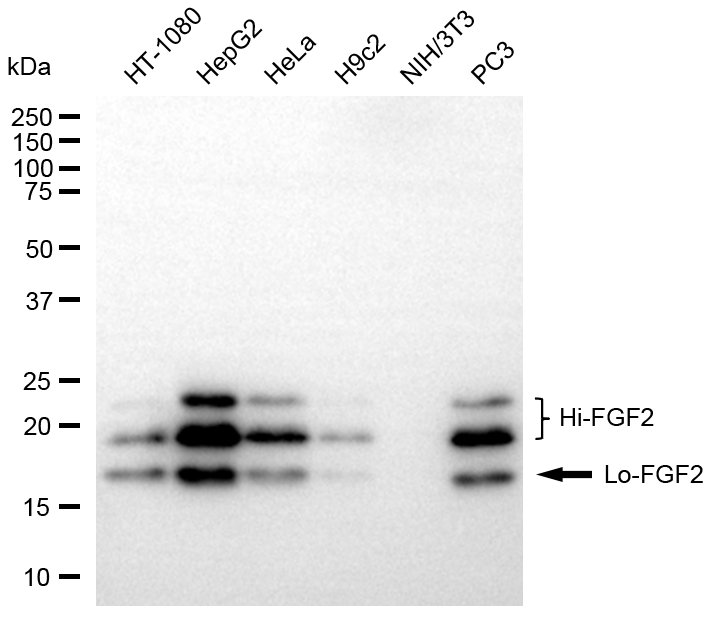
Recombinant Bovine Fibroblast growth factor 2 (FGF2) (Active); CSB-EP008625BOe1
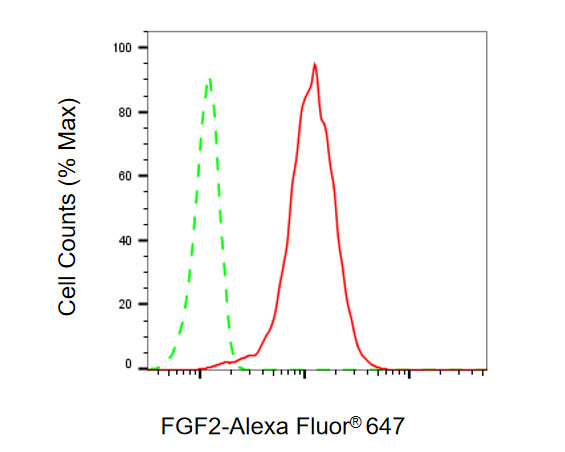
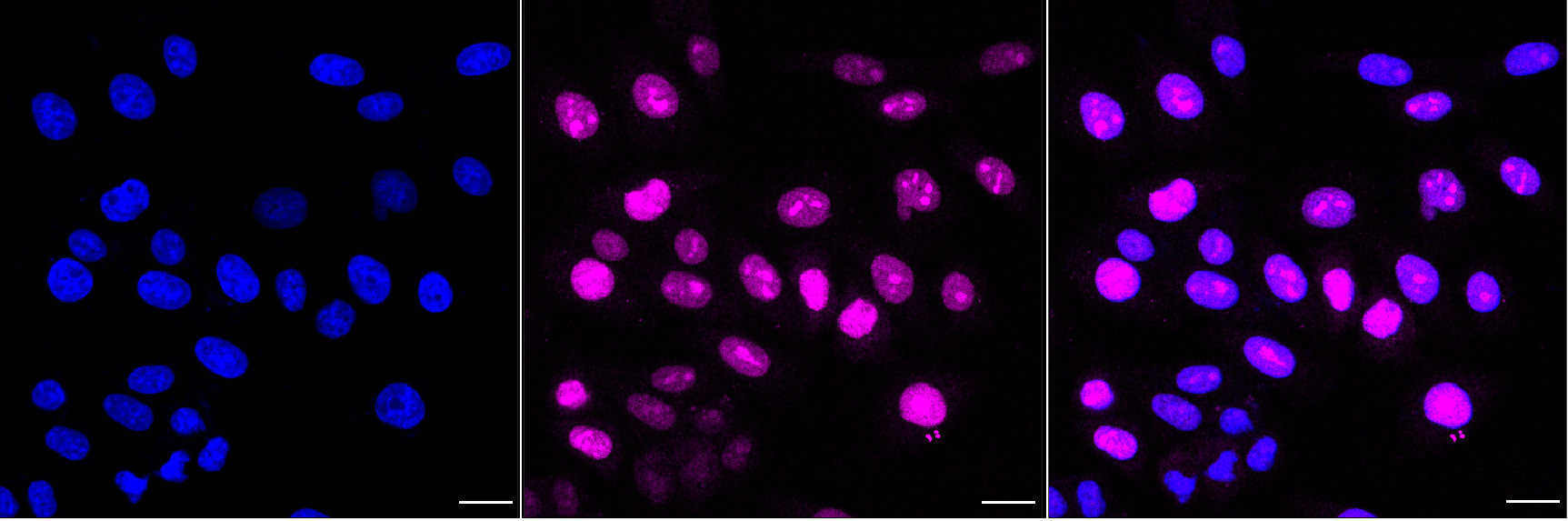
● FGF2 抗體
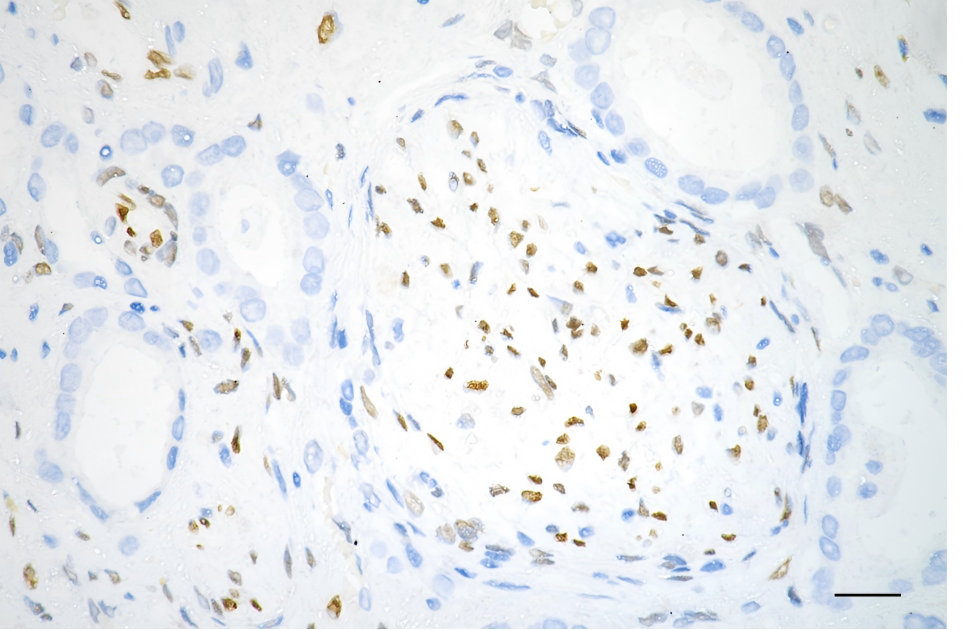
FGF2 Recombinant Monoclonal Antibody; CSB-RA927794A0HU
● FGF2 ELISA 試劑盒
[1] S. Sarabipour, K. Hristova. (2016). Mechanism of FGF receptor dimerization and activation.
[2] Yuga Okada, Akihiro Eguchi, Daisuke Kuroda, Kouhei Tsumoto, Ryosuke Ueki, Shinsuke Sando. (2025). Developing FGF2 Mutants with Selectively Reduced Heparan Sulfate Affinity to Explore Their Impact on FGFR1 Signaling.
[3] W. Chan, K. Howe, James M. Clegg, S. Guimond, D. Price, J. Turnbull, T. Pratt. (2015). 2-O Heparan Sulfate Sulfation by Hs2st Is Required for Erk/Mapk Signalling Activation at the Mid-Gestational Mouse Telencephalic Midline.
[4] Dasol Han, S. Lee, Mookwang Kwon, Hogyun Noh, J. H. Lee, Y. Yoon, J. Cho, K. Yoon. (2020). YAP enhances FGF2-dependent neural stem cell proliferation by induction of fibroblast growth factor receptor expression.
[5] Sebastián Giulianelli, Marina Riggio, Tomás Guillardoy, C. Pérez Pi?ero, M. Gorostiaga, G. Sequeira, Gabriela Pataccini, M. F. Abascal, Maria F. Toledo, B. Jacobsen, Ana C. L. Guerreiro, António S. Barros, V. Novaro, F. Monteiro, Francisco Amado, H. Gass, M. Abba, L. Helguero, C. Lanari. (2019). FGF2 induces breast cancer growth through ligand?independent activation and recruitment of ERα and PRBΔ4 isoform to MYC regulatory sequences.
[6] Bin Zeng, Guosheng Lin, X. Ren, Yan Zhang, Honglei Chen. (2010). Over-expression of HO-1 on mesenchymal stem cells promotes angiogenesis and improves myocardial function in infarcted myocardium.
[7] Xiaotang Wang, W. Fan, Na Li, Yan Ma, Mudi Yao, Guoqing Wang, Siyuan He, Wanqian Li, J. Tan, Qingjun Lu, Shengping Hou. (2021). YY1 lactylation in microglia promotes angiogenesis through transcription activation-mediated upregulation of FGF2.
[8] P. Sun, Feifei Ma, Yang Xu, Chao Zhou, R. A. Stetler, K. Yin. (2021). Genetic deletion of endothelial microRNA-15a/16-1 promotes cerebral angiogenesis and neurological recovery in ischemic stroke through Src signaling pathway.
[9] Lushu Chen, Yuan Cao, Yaming Shen, Huan Li, Rong Ye, Jin Yao. (2023). Downregulation of PIK3IP1 in retinal microglia promotes retinal pathological neovascularization via PI3K-AKT pathway activation.
[10] A. Fouda, Zhimin Xu, J. Suwanpradid, M. Rojas, E. Shosha, Tahira Lemtalsi, C. Patel, Ji Xing, Sumbal Zaidi, W. Zhi, Brain K Stansfield, P. Cheng, S. Narayanan, R. Caldwell, R. Caldwell. (2022). Targeting proliferative retinopathy: Arginase 1 limits vitreoretinal neovascularization and promotes angiogenic repair.
[11] Zijie Wang, Chuying Chen, Liying Sun, Mei He, Ting Huang, Jiji Zheng, Jiayuan Wu. (2024). Fibroblast growth factor 2 promotes osteo/odontogenic differentiation in stem cells from the apical papilla by inhibiting PI3K/AKT pathway.
[12] Zhonghao Wang, Yexin Wang, Yu Tang, Xiaoyan Guo, Qize Gao, Yiming Shao, Jingxuan Wang, Ronghua Tian, Yingxu Shi. (2025). Sodium Benzoate Inhibits Osteoblast Differentiation and Accelerates Bone Loss by Regulating the FGF2/p38/RUNX2 Pathway.
[13] Ok-Jin Park, Hyun-Jung Kim, K. Woo, J. Baek, H. Ryoo. (2009). FGF2-activated ERK Mitogen-activated Protein Kinase Enhances Runx2 Acetylation and Stabilization.
[14] Yang Jiao, Jing Xu, Bingtan Song, Ailing Wu, L. Pan, Ying Xu, Fenghao Geng, Xiaoqian Li, Congzhao Zhao, Min Hong, Xuanyu Meng, Judong Luo, Peng Liu, Ming Li, Wei Zhu, Jianping Cao, Shuyu Zhang. (2022). Interferon regulatory factor 1‐triggered free ubiquitin protects the intestines against radiation‐induced injury via CXCR4/FGF2 signaling.
[15] Mengwei Wang, Yuanmeng Sun, Yanzhi Zhao, Xinyi Jiang, Teng Wang, Junye Xie, Xiuling Yu, Shujun Guo, Yibo Zhang, Xiaojia Chen, An Hong. (2025). An FGF2-Derived Short Peptide Attenuates Bleomycin-Induced Pulmonary Fibrosis by Inhibiting Collagen Deposition and Epithelial--Mesenchymal Transition via the FGFR/MAPK Signaling Pathway.
[16] R. Harris, A. Adeyemo, J. Redfield, Cha-hua Huang, Min Jiang, Xiaoping Ren, W. Jones, Jack Rubinstein, J. J. Schultz. (2016). Long‐term Biological Functions of Fibroblast Growth Factor‐2 (FGF2) in Mouse Hearts Following Myocardial Infarction.
[17] Josh W DiGiacomo, Inês Godet, Michael Trautmann-Rodriguez, Daniele M. Gilkes. (2020). Extracellular Matrix--Bound FGF2 Mediates Estrogen Receptor Signaling and Therapeutic Response in Breast Cancer.
[18] P. Têtu, J. Delyon, J. André, Coralie Reger de Moura, Malak Sabbah, G. Ghanem, M. Battistella, S. Mourah, C. Lebbé, N. Dumaz. (2020). FGF2 Induces Resistance to Nilotinib through MAPK Pathway Activation in KIT Mutated Melanoma.
[19] Yu Zhang, Yan Wang, Di Zhou, Kai Wang, Xu Wang, Xiang Wang, Yang Jiang, Min Zhao, Rutong Yu, Xiuping Zhou. (2021). Radiation-induced YAP activation confers glioma radioresistance via promoting FGF2 transcription and DNA damage repair.
[20] Iwona Bednarz-Misa, Paulina Fortuna, D. Diakowska, Natalia Jamrozik, M. Krzystek-Korpacka. (2020). Distinct Local and Systemic Molecular Signatures in the Esophageal and Gastric Cancers: Possible Therapy Targets and Biomarkers for Gastric Cancer.
[21] Z. Alayash, S. Baumeister, B. Holtfreter, Thomas Kocher, H. Baurecht, B. Ehmke, D. Hagenfeld, S. L. Reckelkamm, M. Nolde. (2025). Proteome-Guided Drug Target Discovery for Periodontitis.















